ПАЛЕОНТОЛОГИЧЕСКИЙ ЖУРНАЛ. 1999, N4, стр. 10З-125
УДК 552.6
Внимание! Копия статьи не вычитана! Сканирование производилось с очень плохого отпечатка!
БАКТЕРИАЛЬНАЯ ПАЛЕОНТОЛОГИЯ
И ИССЛЕДОВАНИЯ УГЛИСТЫХ ХОНДРИТОВ
(c) 1999 г. Л. М. Герасименко*, Е. А. Жегалло**,
С. И. Жмур***, А. Ю. Розанов**, Р. Хувер****
'∙' Институт микробиологии РАН
** Палеонтологический институт РАН
*** Институт литосферы РАН
**** Space Sciences Laboratory. George C. Marshal Space Flight Center, Huntsville. Alabama. USA
Поступила в редакцию 25.03.99 г.
Принята к печати 26.03.99 г.
Описаны модельные объекты на земле, на примере которых ведется изучение биоморфных струк-
тур в космическом материале. Приведены данные по таким структурам некоторых метеоритов.
Описаны трудности бактериально-палеонтологического изучения метеоритов.
Последние несколько лет после ряда публика-
ций (McKay et a!.. 1996. 1997: Hoover, 1997; Жмур
и др., 1997) начался по существу новый этап в изу-
чении биоморфных структур в космическом ма-
териале.
Три важных обстоятельства способствовали
широкому резонансу этих работ. Первое - это то.
что сходные результаты были получены одно-
временно как в России, так и в США. Второе, что
к этому времени широко развернулись работы по
бактериальной палеонтологии (Розанов, Завар-
зин. 1997) или геомикробиологин (Banfield, Neal-
son. 1998). создавшие реальную основу для более
убедительных рассуждений о биоморфах в метео-
ритах, и третье - это широкое развитие работ по
экспериментальной фоссилизашш. ясно показав-
шее. что микроорганизмы могут легко сохра-
няться в ископаемом состоянии, поскольку при
определенных условиях их фоссилизация занима-
ет лишь считанные часы.
Но вернемся сначала к истории.
ИСТОРИЯ ИЗУЧЕНИЯ БИОМОРФНЫХ
СТРУКТУР МЕТЕОРИТОВ
В 1806году.Т.Дж.Тенард(Тпепагс1. 1806) обна-
ружил необыкновенно высокое содержание угле-
рода (2,5%) в каменном метеорите, который упал
15 марта 1806 г. в Алаисе (Франция). Метеорит из
Алаиса, первый известный углистый хондрит,
подвергся интенсивному изучению. В 1834 г.
шведский химик Дж. Берцелпус изучил образец
Аланса (Berzelius, 1834". Он был удивлен, когда
обнаружил воду в этом образце, и почти уверил-
ся, что это не образец метеорита, поскольку он
был убежден, что вода в метеоритах не сохраня-
ется. Когда он узнал, что Алаис действительно содержит воду, он продолжил изучение и устано-
вил, что этот метеорит состоит главным образом
из глинисто-подобных минералов. Берцелиус
также отметил наличие органического вещества
в Алаисе. но грубые аналитические методы 19-го
столетня не позволили ему выяснить, какие имен-
но органические вещества присутствовали в ме-
теорите. Однако, он отметил сходство углпстого
вещества Алаиса с земным биологическим мате-
риалом.
14 мая 1864 г. более, чем 20 черных камней
(некоторые весом более 2 кг) упали внутри эл-
липса длиной 18 км. вытянутого с запада на
восток (Daubree, 1864: Leyineri. 1864). Внутри него
оказались французские деревни Ноик и Оргэй.
Сразу после падения деревенские жители собрали
иссиня-черные камни (многие полностью покры-
тые коркой). Метеорит Оргэй был немедленно
подвергнут тщательному химическому и минера-
логическому анализу Клоэцом и Писани (Cloez.
1864: Pisani, 1864). Было установлено, что он со-
стоит из мягкого, черного горючего материала, с
солями аммония, гумусом, магнетитом, кремние-
вой кислотой. Силикатные минералы скорее все-
го представлены серпентином. Было обнаруже-
но. что минералы метеорита Оргэй напоминают
глины и минералы со структурой, напоминающей
пирокластические осадки, похожие на земные
пепловые потоки, которые сформированы вул-
каническим пеплом при осаждении в воду. Одна-
ко состав метеорита Оргэй совершенно отличен.
поскольку вулканический пепел обычно содер-
жит меньше магния и больше кремния, алюминия
и окислов кальция, чем в метеоритах.
Содержание углерода в обломках метеорита
Оргэй было таким высоким, что поначалу это
рассматривалось как следствие загрязнения зем иым веществом. Соединения углерода в углпстых
хондритах представлены главным образом поли-
меро-подобными материалами, органическими
соединениями (сходными с торфом или лигни-
том). которые не растворяются обычными рас-
творителями. Соединения углерода также обна-
ружены в форме алифатических и ароматических
углеводородов, расемических и хиральных ами-
нокислот. пуриновых и пиримидиновых основа-
ний. жирных кислот и липидов. порфиринов. а
также представлены широким спектром сложной
органики, которая не вписывается в рамки зем-
ной биохимии. Надь (Nagy. 19~5) подробно рас-
смотрел углистые метеориты, геохимию нефти и
комплексные полимеры, керогены и углеводоро-
ды. содержащиеся в углистых хондритах. Ходж-
сон и Бейкер (Hodgson. Baker. 1964. 1969) обнару-
жили порфириновые пигменты в метеорите Ор-
гэй. которые спектрально схожи с земными
осадочными породами. Они обнаружили зоны аб-
сорбции, похожие на ванадпловые порфирнны
(встречающиеся в угле. нефти, и нефтяных слан-
цах). Нефтяники рассматривают ванадиловые
порфнрины как важные показатели биогенного
происхождения нефти.
Они также пришли к выводу, что инфракрас-
ные спектры напоминают органику в древних
земных осадочных породах (например посидони-
евые сланцы). В метеоритах отсутствуют хлори-
ны (производные порфнринов. в которых кольца
пиррола имеют водородные дополнения). Эти
близко связанные пигменты присутствуют прак-
тически во всех земных организмах и должны бы-
ли бы указывать на современное загрязнение
земным веществом. Хлорины медленно преобра-
зуются в порфирины и также отсутствуют в древ-
них земных осадках. Открытие порфиринов в ме-
теорите Оргэй свидетельствует о "весьма вероят-
ном участии живого материала в формировании
метеоритов". Более того. отсутствие хлоринов
свидетельствует о том. что метеоритные порфири-
ны возникают либо в результате пребиотического
синтеза , либо в результате древней микробной
жизни, и не могут быть результатом современно-
го загрязнения земным веществом.
ИСТОРИЯ ИЗУЧЕНИЯ СЛЕДОВ
МИКРОФОССИЛИИ В УГЛИСТЫХ
МЕТЕОРИТАХ
В 1961 г. Б. Надь и его коллеги (Nagy et al.,
1961) сообщили, что масс-спектральный анализ
показал, что органика в метеорите Оргэй имеет
ту же организацию молекулярных фрагментов
(включая фрагменты алифатических и аромати-
ческих соединений углерода), которые найдены в
ископаемом веществе на Земле и в нефти. Клаус
и Надь (Clans. Nagy. 1961) провели микробиоло-
гическое изучение углистых хондритов Оргэй и Ивуна. Это исследование выявило большое число
(-1600/мг) очень маленьких (-10 мкм в диаметре)
тел. которые напоминали ископаемые водорос-
ли. пыльцу и гистрихосферы.Эти тела различной
морфологии были обнаружены во многих углис-
тых хондритах, но они отсутствовали в неуглис-
тых метеоритах Холтрук и Брудерхайм. Изучая
метеориты, эти исследователи нашли и исключи-
ли распознаваемые земные загрязнения в каждом
образце, но при этом пришли к выводу, что в ме-
теоритах присутствует свой комплекс микрофос-
силий. После решительных возражений противни-
ков этих взглядов. Клаус и Надь решили назвать
эти тела (которые, как они считали, являлись ми-
крофосснлиями метеоритов) "организованными
элементами" до тех пор, пока их точная природа
не будет установлена.
Андерс и Фитч (Anders, Fitch. 1962) также об-
наружили эти структуры, но в меньшем количе-
стве и только простой морфологии в их образце
метеорита. Они посчитали сложные структуры
артефактами или загрязнением. Андерс (Anders.
1962а) выступил против биогенной природы ма-
териала. заявив, что ископаемые в метеоритах
"не имеют аналогов" и что биогенную теорию
следует отвергнуть (Anders, 1962b; Fitch. Anders.
1963a.b). Он также заявил, что обнаружил один
образец (из музея Мантауван). который был за-
грязнен (возможно специально) в прошлом веке
(Anders et al.. 1964). Фитч и Андерс были самыми
активными критиками того. что "организован-
ные элементы'' свидетельствуют о присутствии
микрофоссилий в метеоритах. Они предположи-
ли. что эти тела не были биологическими объек-
тами и посчитали все другие биологически-по-
добные тела земными загрязнениями, такими как
"музейная пыль" или "пыльца". Клаус и Надь
(Clans. Nagy. 1961) вполне отдавали себе отчет в
проблеме загрязнения и они очень детально рас-
смотрели ее в своей первой статье. Все определи-
мые земные микробы или частицы были отмече-
ны и рассматривались как земные загрязнения.
Они обнаружили примерно 100 бактерий. 3 диато-
меи. 2 спикулы губок. 1 гранулу крахмала и 16 во-
локон целлюлозы. В препаратах под микроско-
пом. содержащих более 5000 форм. они опреде-
лили "организованные элементы", которые они
выделили из метеоритов Оргэй и Ивуна. Деталь-
ное изучение биологических материалов в почве
и осадочных породах в окрестностях Оргэй (Nagy
et al.. 1962) позволило оценить возможное дейст-
вие заражения земным веществом. Они также об-
наружили идентичные микроструктуры ("орга-
низованные элементы") в метеоритах Тонк и
Алаис. Это показало, что источником данных
структур не было локальное земное загрязнение
(Clans et al.. 1963). Клаус и Надь использовали
стандартные мнкропалеонтологические и пале-
онтологические методы и провели биологичес кое окрашивание этих микроструктур, чтобы
различить органические и минералогические ча-
стицы. Характер окрашивания iNagy et al., 1963b)
и ультрафиолетовые спектры (Nagy et al.. 1963a)
микроструктур, обнаруженных в метеоритах,
привели их к выводу, что "организованные эле-
менты" имеют биологическое происхождение.
Таш (Tasch, 1963, 1964) тщательно изучил явле-
ния загрязнения и отметил, что хотя в углистых
хондритах были обнаружены распознаваемые
земные загрязнения, они не могли объединить
большинство инснтных и очевидно биогенных
микроструктур. Более того, эти формы не смогли
определить высококвалифицированные микро-
палеонтологи. которые исследовали препараты и
проводили свои собственные исследования углис-
тых хондритов.
Возможно, наиболее важное независимое ис-
следование природы и "организованных элемен-
тов" Клауса и Надя было проведено известными
палинологами М. Россигнол-Стрик и Э. Барг-
хурном, ведущими специалистами по древним
микрофоссилиям. Используя стандартные пали-
нологические методы. Россигнол-Стрик и Барг-
хурн обрабатывали внутренние, чистые образцы
из метеорита Оргэй с помощью реактива Шульца
(HNO', и КС10'.) и концентрированной плавиковой
кислоты (HF) в течение одного часа. Они обнару-
жили множество полых сфер и ооидов, мембраны,
"воронковидные" и "грибовидные" структуры и
т.п. (Rossignol-Strick. Barghoorn. 1971). Многие из
этих тел из метеорита Оргэй. как было установ-
лено, являются близкими по размеру, форме и
внешнему облику к микрофоссилиям (Ниго-
niospora sp.) из ганфлинтских кремней.
Эти сферы имеют тот же цвет. плотность и со-
став, как и включающий коричневый органичес-
кий осадок. Более того. они имеют хорошо очер-
ченные стенки (похожие на клеточные стенки) с
гладкими внутренними поверхностями и неров-
ными внешними поверхностями из-за присутст-
вия частиц породы на внешней стороне сфер. Не-
которые из этих стенок было обломаны и созда-
ют впечатление эластичности. Эти органические
тела негативно реагируют на палинологический
краситель шафранин и поэтому не являются
пыльцевыми зернами. Их также нельзя спутать
со спорами грибов или волокнами тканей. Эти
кислото-устойчивые микроструктуры имели
близкий размер и черты с "организованными эле-
ментами", изученными Клаусом и Надем. Барг-
хурн и Россигнол-Стрик сообщили, что "обилие
органических объектов в метеорите Оргэй мо-
жет быть свидетельством биогенного происхож-
дения". Полые сферы метеорита Оргэй также
похожи морфологически на кислотоустойчивые
полые сферы размером 6-20 мкм, которые на-
блюдаются в осадочных породах, подстилающих
серию Фиг Три (серия Онфервахт (Onverwacht) возрастом более 3.2 млрд. лет). Гистограммы
выявили, что распределение по размеру полых
микроструктур в метеорите Оргэй такое же, как
в популяциях мнкрофоссилнй из железистых
кварцитов Ганфлита в Онтарио (Канада) возрас-
том два миллиарда лет. Многие из "организован-
ных элементов" похожи по размеру, форме и
внешнему облику на разнообразные докембрий-
ские микрофорссилии (например. Eosphaera tyieri.
Melasmatosphaera sp., Huroniospora sp.). Хотя тело
метеорита Оргэй содержало жидкую воду. оса-
дочная текстура не наблюдалась. Они отметили.
что "на Земле фоссилии обнаружены только в
осадочных породах" (Rossignol-Strick. Barghoorn.
1971). И поэтому считали, что биогенная природа
должна быть связана с окружающей средой, и по-
скольку метеоритная порода не была осадочной.
то данные формы имели абиогенное происхожде-
ние. В то время микробные сообщества гидротер-
мальных источников, глубинных базальтовых и
гранитных пород и вулканических туфов были
неизвестны, и палеонтологи не ожидали встре-
тить ископаемые в неосадочных породах.
Другие исследователи загадочные микрост-
руктуры в углистых метеоритах относили к мик-
рофоссилиям биологического проихождения. ко-
торые принадлежат данным метеоритам. Они от-
мечали, что эти тела нельзя идентифицировать с
известными земными микроорганизмами. Б. Ти-
мофеев (Timofejev. 1963). который был одним из
родоначальников палинологии, выделил из мете-
орита Мигей (путем воздействия азотной кисло-
ты с последующим цетрифугированием) большое
число нерастворимых в кислоте сферических
форм, которые были очень похожи на докемб-
рийские акритархи. Бриггс и Китто (Briggs. Kitto.
1962) также нашли сложные одноклеточные те-
ла. напоминающие микроорганизмы, в метеори-
те Мокоя. Надь и др. (Nagy et al.. 1963b). использо-
вав микроанализатор, показали, что некоторые
"организованные элементы" были насыщены лн-
монитом, содержащим немного никеля, а другие -
силикатами. Используя НС1 и HF. чтобы раство-
рить минералы, они обнаружили, что большин-
ство оставшихся углистых тел не содержали эле-
метов с атомным номером выше магния. Это они
расценили как доказательво фоссилизации. кото-
рое ясно исключает земное загрязнение, как ис-
точник этих тел. Стаплпн и ВанЛандингхам (Stap-
lin. 1962; VanLandingham. 1965: VanLandmgham et
al.. 1967) также заключили, что эти похожие на
биологические структуры образования были
фоссилизированными микроорганизмами в мете-
оритах. Флюоресцентные исследования метеори-
тов (Alpem, Benkheiri. 1973) показали, что боль-
шая часть органического вещества в метеорите
Оргэй заключается в этих маленьких сферах ди-
аметром примерно 1-10 мкм. Росс (Ross. 1963)
также встретил грибовидные полые трубки с рваными мембранами в самой легкой части мате-
риала метеорита Оргэй. который он выделил с
помощью суспензий кадмия. Чтобы свести к ми-
нимуму возможные загрязнения, в его анализе
применялись стерильные инструменты, которы-
ми вскрывался нетронутый метеорит Оргэй из
Британского музея.
Тан и ВанЛадингхам Пап. VanLadingham. 1967)
получили фотографии с электронного микроско-
па многих кислотоустойчивых тел, которые они
рассматривали как микрофоссплии. которые вхо-
дили в состав метеоритов Оргэй, Ивуна и Тонк.
Использовавшиеся сильные окислители - азот-
ная. соляная и плавиковая кислоты уничтожили
бы карбонатные и кремниевые микрофоссилии,
которые содержатся в большинстве земных сооб-
ществ (диатомовые. силпкофлагеллаты. форамн-
ниферы и кокколиты). Они обнаружили многие
морфологические формы, включая "грибообраз-
ные" структуры с темными под эелектронным
микроскопом "ножками" и более прозрачные,
мембрановидные шляпки со складками. Некото-
рые из этих форм они посчитали очень схожими
по размеру, форме и особенностям изображений
под электронным микроскопом с углистыми на-
ночастицами. которые Пфлюг и Хайнц (Phlug,
Heinz. 1997) обнаружили в барите из формации
Фиг Три возрастом 3.3 млрд. лет. Они также по-
счптали нити с одним закругленным и другим за-
остренным концами, которые содержали цепь не-
прозрачных образовании (вероятно, кристаллов)
вдоль длинной оси. очень сходными со структура-
ми. которые теперь описываются из магнетотак-
тическнх бактерий. Палик (Palik. 1962) обнаружи-
ла и изобразила сложные структуры, похожие на
нитчатые водоросли, в механически разрушен-
ных (не обработанных кислотой) образцах мето-
рита Оргэй. Некоторые нити были загнуты, их
ширина менялась от 3 до 16 мкм. а длина от 40 до
170 мкм. Она описала необыкновенную нить N 2
как "часть нити шириной 4 мкм и длиной 55 мкм.
Она покрыта фолликулом, вершина закруглена".
Эти структуры "немного утонялись к верхушке".
Мы тоже обнаружили удивительно похожие нити
in situ в метеорите Мурчисон i Hoover. 1997). Не-
которые имеют фолликуловидную калиптру в
апикальной части. Палик описала некоторые
формы из метеорита Оргэй. которые были орна-
ментированы. Как она пишет, "на двойной стенке
в разных местах можно наблюдать маленькие
узелки" (Palik. 1962). Мы обнаружили похожие
структуры на полых нитях в метеорите Мурчи-
сон. Некоторые из нитей, описанных Палик, за-
канчиваются расширениями.
Существование внеземных "жизнеподобных"
форм было достаточно широко принято до сере-
дины 60-х годов, и некоторые палеонтологи нача-
ли изучать проблемы систематического положе-
ния внеземных микрофоссилнй (Claus. Nagy, 1962; VanLandingham. 1963). Однако, последовала
интенсивная критика, и противоречия, связанные
с "организованными элементами", были такими
сильными, что к середине 70-х годов большинст-
во исследователей прекратили какое-либо изуче-
ние возможных микрофоссилнй в метеоритах.
Недавно статьи в популярной прессе отнесли все
наблюдения "организованных элементов" в мете-
орите ALH84001 просто к нераспознанной пыль-
це деревьев или "смятым пыльцевым зернам амб-
розии". Это мнение не поддерживается в научной
литературе. Загрязнения спорами и пыльцой бы-
ли обнаружены (Gregory. 1962; Pearson. 1962). но
загрязнения земными веществами не могут объ-
яснить многие предположительно биологические
тела (найденные глубоко в метеоритах разными
исследователями), которые обладают сложной
морфологией, которая неизвестна в микропале-
онтологии. палинологии и микробиологии. Похо-
жие тела были найдены во многих углистых хон-
дритах (Оргэй. Тонк, Ивуна, Алаис. Мурчисон.
Муррей, Мигей и Мокоя). но не найдены в не\тли-
стых метеоритах (Холбрук. Брудерхайм. Индарч.
Пис Ривер).
В 1966 г. Х.С. Урей (Urey. 1966) (лауреат Нобе-
левской премии по химии 1934 г.) сделал обзор
доказательств биологических материалов в мете-
оритах. Он отметил, что в метеоритах встречают-
ся органические субстанции, которые близко на-
поминают таковые из древних земных пород, что
органика, встреченная в углистых хондритах. не
напоминает ту, которая присутствует в современ-
ных загрязнениях. Урей отмечал: "...некоторые
вещества в метеоритах, если бы их нашли в зем-
ных объектах, несомненно бы считались биоген-
ными". Он также заметил: "... те из нас. кто в те-
чение нескольких лет работали с метеоритами.
были уверены, что там не могло быть остатков
никаких живых организмов. Если бы метеориты
имели состав осадочных пород Земли, это не вы-
звало бы большого удивления".
МОДЕЛЬНЫЕ ОБЪЕКТЫ .
Начало исследований метеоритов авторами
статьи определилось опытом исследований древ-
них высокоуглеродистых пород и фосфоритов, а
также опытами по лабораторной фоссилизации
цианобактерий (Жмур и др., 1989. 1993. 1995;
Жмур, 1993; Розанов, Жегалло. 1989: Крылов.
Орлеанский.1988: Герасименко и др.. 19^4. 1996.
1998). Именно эти работы сформировали новое
направление - "бактериальную палеонтологию"
(Розанов, Заварзин, 1997). Три обстоятельства
имели решающее значение:
Первое - это осознание того. что микробы, и
особенно цианобактериальные маты. прекрасно
сохраняются в ископаемом состоянии и могут
быть обнаружены практически в любых типах пород, при этом. правда. длительная история изу-
чения древних ископаемых микробов в основном
сводилась к микроскопическом} изучению крем-
нистых пород и окремненных участков стромато-
литов (Shopf. Klein (eels), 1992;" Knoll. Walter, 1992:
Сергеев и др.. 1998):
Второе ~ это осознание того. что процесс фос-
силизации происходит практически мгновенно, в
считанные часы: и
Третье - это осознание того. что все высокоуг-
леродистые породы так или иначе связаны с бак-
териальной активностью, что и навело на мысль
о возможности присутствия псевдоморфоз по ми-
кроорганизмам в углистых хондритах.
Кроме указанных обстоятельств, огромное
значение имело длительное изучение бактери-
альных ископаемых в древних фосфоритах, поз-
волившее проследить разную степень сохраннос-
ти цианобактерий (Розанов, Жегалло, 1989; Zhe-
gallo et al., 1999). Особо важно при этом было
постоянное сравнение с современными материа-
лами.
Таким образом, накопленный опыт подсказал.
что основными модельными объектами должны
быть фосфориты, высокоуглеродистые породы и
современные цианобактерии для дальнейших
опытов по закономерностям фоссилизации.
СОВРЕМЕННЫЕ ЦИАНОБАКТЕРИИ
И ИХ ФОССИЛНЗАЦИЯ
Цианобактерии - древнейшие фотосинтезиру-
ющие организмы, жизнедеятельность которых
разгрузила древнюю атмосферу Земли от угле-
кислоты и обеспечила ее кислородом. Именно
цианобактерии вместе с сопутствующими им бак-
териями более, чем на 3 млрд. лет стали полными
властелинами Земли и в значительной степени
определили ход таких важных геологических со-
бытий. как накопление многих осадочных пород
и полезных ископаемых. Созданные этими мик-
роорганизмами сообщества, имеющие тесные ме-
таболические связи, оказались удивительно ус-
тойчивыми на протяжении всей истории Земли.
Правда, более высокоорганизованные конку-
ренты постепенно вытеснили их с широких мор-
ских просторов эпиконтинентальных бассейнов в
экологические ниши в основном с экстремальны-
ми условиями, такими как гиперсоленые лагуны.
вулканические области и др. II в этих местах ми-
кробные сообщества сохраняются до настоящих
дней.
Термофильные сообщества мы можем наблю-
дать в кальдере вулкана Узон на Камчатке и на о.
Кунашир. галофильные сообщества - в лагунах
оз. Сиваш в Крыму.
Основу сообществ (матов) обычно составляют
осциллаториевые цианобактерии. В термофильных матах при температуре до 4()╟С это различ-
ные виды Phormidium, при более высоких темпе-
ратурах доминирует плеоморфный Mastigocladus
laminosus - на Узоне или Synechococcus elongatus -
на Кунашире.
Все маты имеют слоистую структуру, п каж-
дом из слоев развиваются определенные виды
микроорганизмов. Цианобактерии занимают
верхний слой. Из фототрофных бактерий, разви-
тие которых приурочено к слою цианобактерий.
следует отметить Chloroflexus ;iiirantiacus для тер-
мофильных матов и Thiocap.sa или Ectothiorno-
dospira - для галофильных. Эти бактерии в основ-
ном осуществляют органотрофный рост. окисляя
продукты жизнедеятельности цианобактернй. хо-
тя могут окислять сероводород и участвовать в
цикле серы.
В гидротермах часто обнаруживаются мине-
ральные постройки с отложением травертина.
кремнезема, напоминающие строматолнты. Это
позволяет при изучении процесса минерализации
в природе, а также на модельных опытах в лабо-
ратории. получить представление о деятельности
цианобактерий в геологическом прошлом.
В термальных источниках с высоким содержа-
нием кремнезема некоторые участки сообщества
минерализованы. В таких окремнелых корках
были обнаружены минерализованные остатки ми-
кроорганизмов - современные мнкрофосснлии.
Сопоставляя различные фоссилии. И.Н. Крылов и
др. (1989) проследили разные стадии окремнения.
Картина, представленная ими. заключается в сле-
дующем. Сначала кремнезем проникает внутрь
клетки и осаждается в виде глобул на клеточных
стенках и перегородках. На следующей стадии
отдельные глобулы сливаются в сплошную скор-
лупку, окружающую клетку. Наружная оболочка
такой скорлупки гладкая и повторяет форму кле-
точной оболочки, внутренняя - глобулярная.
В дальнейшем опаловые глобулы заполняют пу-
стоты между нитями и внутри них до тех пор. по-
ка не образуется сплошная окремнелая порода
(рис. 1, а). Была отмечена одна особенность: ок-
ремнение шло только в мертвых клетках. Под-
тверждение этого было получено в лабораторных
опытах. Кремнезем замещал только оболочки и
перегородки нитей, находящихся на разных ста-
диях посмертного разложения (рис. 1, о). Таким
образом, живая клетка имеет механизмы защиты
от высоких концентраций кремния, в первую оче-
редь образуя толстый слизистый чехол (рис. 1, el.
Проникновение кремния возможно только после
гибели клетки и разрушения защитных свойств
клеточных мембран.
И в термофильных, и в галофильных матах ча-
сто прослеживаются карбонатные включения.
представленные арагонитом и кальцитом. Обна-
руженные в этих областях цианобактерии также

Рис. 1. и - окремнецис нитей \b-:.;:ocladus laminosum в природном термофильном мате: б, в - процесс окремененчя в
лабораторной культуре Phormid:-:'i laminosum: n - мертвые клетки, о - живые клетки: г - карбонатп ;ация и культуре
Phonnidium laminosum. живые тр;'\омы и образование слизистого чехла с отдельными глобулами СаСО^.
карбонатизированы. Строгая корреляция содер-
жания карбоната Са++ с хлорофиллом цианобак-
терий в мате (Заварзин. 1989 говорит о том. что
одной из причин, безусловно не единственной, яв-
ляется подщелачивание срезы в результате ак-
тивного фотосинтеза цианобактерий. В лабора-
торых опытах показано, что выпадение карбона-
та коррелируется с состоянием культуры и имеет
максимальное значение при оптимальных усло-
виях фотосинтеза (Некрасова и др., 1983). Увели-
чение содержания карбоната Са+ + в среде вело к
морфологическим изменениям, которые на пер-
вых этапах выражались в образовании слизисто-
го чехла. В дальнейшем на чехле оседали глобу-
лы карбоната, которые в последствии мннерали-
зовали весь трихом (рис. 1. г: 2. а).
Находки современных фоссилизированных ос-
татков микроорганизмов в окремненнных и
обызвествленных цианобактериальных матах,
морфологически похожих на древние микрофос-
силии, заставили обратить внимание на формы
сохранности цианобактерий в разных участках
колоний и сообществ в лабораторных условиях
(Герасименко. Крылов. 1983'. Проведенные опы-
ты позволили сделать вывод, что отмеченное разнообразие кремнистых микрофоссилий Узона
определяется не только видовым разнообразием
цианобактерий, но и морфологическим разнооб-
разием клеток (например Mastigocladus lamino-
sus). а также разной степенью посмертной дегра-
дации клеток и нитей.
При рассмотрении вопроса минерализации цп-
анобактерий особенно интересен процесс их фос-
фатизации. Неоспоримое сходство органических
остатков, встречающихся в древних фосфоритах.
с современными (Zhegallo et al.. 1998, 1999). чаше
всего с цианобактериальными. а также сходное
строение слойков древних фосфоритов и совре-
менных цианобактернальных матов, позволяют
ставить вопрос о преимущественной роли именно
цианобактерий в древнем фосфатонакоплении.
При изучении фосфатизации в качестве тест-
объекта была выбрана галофильная цианобакте-
рия Microcoleus chthonoplastes, выделенная из соле-
ных лагун оз. Сиваш. Microcoleus chthonoplastes -
типичный бентосный организм, массовая форма
цианобактериальных матов гиперсоленых водо-
емов аридной зоны (Cohenetal.. 1984; Cohen. Rosen-
berg. 1989; Гераснменко и др., 1989; Stal, Caumette.
1994). Проведенные эксперименты показали, что

Рис . 2. ч - карбонатизация в культ', ре Phormn-iium laminosum, выползание трихома из минерального чехла: п-г - раз-
личные этапы фосфатизации нитей Microcoleus chlhunoplastes: о-трубочки апатита (после добавления NaF): н - пус-
той минеральный чехол; г - минерализация трихома.
М. chthonoplastes способен расти в широком диа-
пазоне концентрации неорганического фосфора.
Независимо от концентрации, активный вы-
нос фосфора из среды отмечался в первые 3 часа,
т.е. в лаг-фазу, и заканчивался к 1-ым суткам (Ти-
хомирова, Орлеанский. 1994; Герасименко и др.,
1996,1998).
Проведенные опыты показали, что не весь
фосфор, поглощенный клеткой в первые мину-
ты, проходит внутрь клетки: большая часть его
адсорбируется на поверхности. И чем выше кон-
центрация фосфора в среде, тем выше процесс
адсорбции.
Известно, что поступление фосфора в клетку
происходит при активном его транспорте через
мембрану (Кулаев, 1975). и он накапливается там
в большей концентрации, чем во внешней среде,
и большей концентрацией, чем требуется клетке
для ее метаболизма. Этот "лишний" фосфор на-
ходится в клетке в виде легко растворимых поли-
фосфатных волютииовых гранул. При разложе-
нии такой биомассы происходит выход фосфора в окружающую среду в количествах, достаточ-
ных для начала минерализации.
У М. chthonoplastes активный транспорт проис-
ходит с максимальной скоростью в первые 3 мин.
и равен 17 мкг/мин '^Р г. а.с.б. (абсолютно сухой
биомассы). Через 3 суток скорость снижается на
два порядка.
При разном содержании фосфора в среде на-
копление его в клетках также различно и возра-
стает пропорционально повышению его концен-
трации в среде, но до определенного предела
(Гончарова, Герасименко,1993).
Повышение концентрации Рненрг ведет к мор-
фологическим изменениям. На поверхности кле-
ток появляется слизистый чехол, толщина кото-
рого иногда вдвое превышает толщину нитей.
Ослизнение у цианобактерий, как и у многих дру-
гих микроорганизмов. - это ответ на неблагопри-
ятные условия среды. Таким образом слизистый
чехол выполняет защитную функцию. Он состо-
ит из аморфной массы, в которой заключены мик-
рофибриллы полисахаридов. Они могут служить
центрами минерализации, образуя минеральныемикротрубочки вокруг ните;'' цианобактерий (рис. 2. б). Выделяющаяся цианобактериями и
другими микроорганизмами слпзь (глнкокаликс)
сохраняется и может минерализоваться. Обнару-
живаемый в древних отложениях фоссилизиро-
ванный гликокаликс является одним из доказа-
тельств жизнедеятельности микроорганизмов.
При дальнейшем повышении концентрации
фосфора на поверхности клетк;; в слизистом чех-
ле возникают отдельные глооулы, которые в
дальнейшем объединяются в бляшки, муфты, об-
разуя в конце концов сплошной минеральный че-
хол (рис.2,в).
Осаждающийся фосфат замещает только сли-
зистый чехол, не трогая сами трихомы. Клетки
при этом остаются жизнеспособными, и в ряде
случаев наблюдалось, как из таких минеральных
чехлов выползают трихомы, оставляя полую
трубку (рис. 2, в). При увеличении концентрации
выше 2 мМ начинается гибель культуры, и проис-
ходит минерализация самих трпхомов (рис. 2, г).
Фосфат кальция замещает и трихомы и разло-
жившуюся детритную массу и. кроме того, осаж-
дается химическим путем. Наступает момент,
когда вся масса представляет собой агрегат, в ко-
тором на фоне общей минеральной массы хорошо
видны отдельные полые трубки, оставшиеся после
выхода трихомов. Картина очень напоминает фо-
сфоритовые желвачки из Хуосугульского фос-
форитового месторождения (рис. 4, в, г: 5, а, б}.
Рентгеноструктурныи анализ состава 10 мине-
ральных чехлов, образующихся вокруг нитей
цианобактерий. и осадка, образованного при де-
градации массы цианобактерин. показал, что по-
лучаются плохо раскрнсталлизованные фосфат-
ные минералы группы апатита с основным дифрак-
ционным максимумом, близким к франколиту
(Герасименко и др., 1994).
Для получения раскристаллизованного апати-
та в лабораторных условиях з среду добавляли
NaF. поскольку F является изоморфным элемен-
том решетки апатита (Goncharova et al., 1993).
В этом случае наблюдалось моментальное выпа-
дение осадка, а изучение под микроскопом пока-
зало наличие минеральных чехлов вокруг нитей
цианобактерий. состоящих из микроскопических
трубочек, сложенных сформированными глобу-
лами микронного размера (рис. 2. о). Микрозондо-
вый анализ показал, что в состав трубочек кроме
Са и Р входит F. т.е. они сложены фторкарбоната-
патитом.
Большие скорости поглощения фосфора
именно в первые минуты объясняют хорошую
сохранность морфологии цианобактерий в древ-
них фосфоритах, что указывает на чрезвычайно
быстрый процесс фосфатизации, когда клетки
еще не успевают разложиться.
ЧЕРНЫЕ СЛАНЦЫ.
ВЫСОКОУГЛЕРОДИСТЫЕ ПОРОДЫ.
ДОМАНИКОИДЫ И Т.Д.
Проведенные исследования значительного ко-
личества объектов высокоуглеродистых пород
(горючих и черных сланцев) Русской и Сибирской
платформ и прилегающих к ним складчатых об-
ластей из широкого возрастного диапазона, начи-
ная с раннего протерозоя и кончая неогеном, по-
казали несомненное участие в их формировании
цианобактериальных бентосных сообществ (ма-
тов). Был выявлен достаточно обширный ком-
плекс литифицированных остатков микробных
организмов, составлявших эти сообщества.
Исследования раннепротерозойских шунгитов
Карелии (Жмур и др., 1993) выявили в стратифи-
цированных их разностях многочисленные остат-
ки коккоидных колониальных бактерий, кото-
рые по своей морфологии близки к современным
бентосным формам типа Gloeocapsa (=Pleurocap-
sa), формирующих в солоноватых и соленых во-
доемах постулярный (пузырчатый) мат в зоне ли-
торали (Bauld. 1984). Представленный на рис. 3, а
литифицированный остаток макроколонни дан-
ного организма демонстрирует все основные
элементы его строения - колониальность, обус-
ловленную вхождением в него целого ряда мик-
роколоний и даже входящие в состав последних
отдельные клетки. В другом раннепротерозой-
ском объекте - черных сланцах Курской магнит-
ной аномалии, были установлены (Жмур. Горлен-
ко, 1989) многотрихомные микроорганизмы типа
современной цианобактерий Microcoleus. харак-
терной для гладких аэробных микробных матов.
развивающихся в мелководных прибрежно-мор-
ских условиях, лагунах, заливах и т.п. (Bauld.
1984).
И в шунгитах. и в черных сланцах. Курской
магнитной аномалии были встречены также ли-
тифицированные остатки одиночных нитей типа
современных Oscillatoria и Phormidium в виде
только одних сохранившихся чехлов или полно-
стью литифицированных. в которых иногда разли-
чается и клеточное строение (рис. 3, б, в). Помимо
остатков микроорганизмов и в тех и в других со-
хранились и следы их жизнедеятельности - гли-
кокаликс. На рис. 3, г представлены его минера-
лизованные остатки из шунгитов, как наиболее
полно передающие характер его построения.
практически ничем не отличимые от фиксируе-
мых в современных матах, когда клеточный ма-
териал полностью деградирован. Такой пример
рассмотрен Крумбейном и Свартом в мате Га-
виш-Себкхи (Krumbein, Swart, 1983). В тех случа-
ях, когда клеточный материал не деградирован.
он может настолько плотно покрывать организмы
мата, что их бывает трудно идентифицировать.

Рис. 3. Литифицированные микроорганизмы из шунгитов: а - колониальная коккоидная цианобактерия типа Gloeo-
capsa: о. н - в различной степени сохраненные остатки нитчатых микроорганизмов типа Phormidium, Oscillatoria: 5-
полностыо литифицированная нить. " - нить с сохранившейся клеточной фрагментацией: г - литифицированныи гли-
кокаликс.
Напомним, что рифей стал временем щиро-
чайшего развития разнообразных карбонатных
строматолитов. обязанных своим происхождени-
ем прежде всего цпанобактерпям (Крылов, 1963,
1975; Комар и др.. 1965; Комар. 1966: Семихатов,
1974: Серебряков. 1975: Семихатов. Раабен, 1993.
1994. 1996). Это было и время широкого развития
углеродистых пород, в формировании которых
принимал участие цианобактериальный мат. Бу-
дучи основной биотоп и обладая способностью в
той или иной степени прижизненной и посмерт-
ной литификацией чехлов и капсул, а также спо-
собностью изменять щелочно-кислотное равнове-
сие водной среды в результате фотоспнтетической
деятельности, это сообщество на докембрийском
этапе, по крайней мере, на большей его части,
осуществляло регуляцию химического состава
вод, убирая из них и переводя в осадок такие эле-
менты как Si. Ca, Al. Fe, Р. Мп.
Наличие мелководных платформенных эпи -
континентальных морей способствовало широ-
кому развитию микробиальных сообществ и в па-
леозое.
Появление высокоорганизованных форм в мо-
рях сказалось на сохранении жизнеустойчивостп
бентосных микробиальных систем, т.к. они брали
на себя значительную часть по переработке по-
ступающего в бассейн терригенного материала и
по очищению вод от избытка содержащихся в
нем различных элементов.
В формировании высокоуглеродистых пород
палеозоя ведущую роль играло коккоидное циа-
нобактериальное бентосное сообщество, кото-
рое формировало характерные для этого време-
ни так называемые доманикоиды - кремнисто-
карбонатные битуминозные горючие сланцы с
очень незначительным количеством глинистого
материала, которые рассматриваются как нефте-
материнские породы. Образованые с помощью
коккоидного мата, они вместе с тем содержат и
нитчатые микроорганизмы. На рис. 4, д видны в
той или иной степени деструктированные кокко-
ндные организмы типа Gloeocapsa (куонамские
сланцы). В доманиковых сланцах обнаружены'
тонкие (0.2-0.3 мк в диаметре) нити. которые
можно отнести к представителям Chloroflexus
фис. 4, б).

Рис. 4. а, б - литифицированные остатки микробных организмов из куонамских и доманиковых сланцев палеозоя: а -
деструктированные коккоидные организмы типа Gloeocapsa (куонамские сланцы), б- предположительно остатки зе-
леной бактерии типа Chloroflexi;- доманиковые сланцы); в, г- хубсугульские фосфориты, фрагменты фосфатчзиро-
ванного цианобактериального \'л-а.
Следует заметить, что исследования карбона-
тов. содержащих в топ пли иной стелен?! глинис-
тый материал, и глинистых сланцев дали также
положительный результат. В глинистых породах
венда Восточно-Европейской платформы, а так-
же известняках кембрия Южной Австралии были
обнаружены легко различимые коккоидные и
нитчатые формы. Это позволяет надеяться, что
фоссилизированные микроорганизмы будут об-
наруживаться во всех осадочных породах, и что
модельными б\'д\'т. кроме высокоуглеродистых
пород и фосфоритов, и другие типы осадочных
пород.
ФОСФОРИТЫ
Исследованию биогенных аспектов образова-
ния древних (докембрнй-кембрий) фосфоритов
посвящена огромная литература. Долгое время
считалось, что фосфориты наиболее крупных ме-
сторождений образуются хемогенным путем (Ильин, 1973. 1990). Одно из наиболее крупных
месторождений - Хубсугульское в Северной
Монголии считалось эталоном хемогенного фос-
фатонакопления (Ильин. 1973, ,1990; Яншин.
Жарков, 1986). Многие исследователи подозрева-
ли однако, что так называемые хемогенные фос-
фориты на самом деле образуются каким-то био-
генным путем (Еганов. 1988: Школьник. 1989: и
др.). Наиболее отчетливый результат был полу-
чен в 1987 г. при электронно-микроскопическом
изучении фосфоритов Хубсугула (Розанов. Же-
галло, 1989). Первоначально изучение фосфори-
тов приводилось в связи с проблемой становления
древних скелетных организмов, когда было заме-
чено, что крупнейшее фосфоритовое месторож-
дение имеет тот же томмотский возраст, что п
многочисленные фосфатные ископаемые, так
называемые SSF (Лувсанданзан. Розанов, 1986:
Розанов, Жегалло, 1989). Поэтому электронно-
микроскопическое исследование фосфоритов
было направлено вначале на поиск органических

Рис. 5. Фосфориты Хубсугульско-с месторождения: а- фосфатный онколит: б-фосфатный микрожелвак: в - веро-
ятно, аморфные глобули апатита;; - крупные плоские дискоидальные кристаллы апатита.
остатков в фосфоритах. Однако очень скоро мы
смогли убедиться, что все фосфориты сложены
фоссилизированными цианобактериальными ма-
тами. состоящими из собственно цианобактерий
(ЦБ) и комплекса других микробов и прежде все-
го пурпурных бактерий (рис. -. б. г: 5, а-г). Со-
хранность ЦБ была совершенно удивительна и
это дало возможность предположить, что ско-
рость фоссилизации была очень большой, что и
подтвердилось позднее в экспериментах (см. вы-
ше). Подробное изложение фактического мате-
риала по Хубсугульскому месторождению с мно-
гочисленными фотографиями древних и совре-
менных цианобактерий и других микробов
опубликовано в специальном выпуске (Zhegallo et
al.. 1999), и здесь мы ограничимся лишь приведе-
нием некоторых фотографии и некоторыми за-
мечаниями об условиях, в которых формирова-
лись фосфориты Хубсугула. Проанализирован-
ные известные данные показывают (Розанов.
1992; Zhegallo et al., 1999). что Хубсугульский бас-
сейн находился в прнэкваториальной зоне, был
мелководен, временами замыкался и образовы-
вался огромный залив. Временами, в краткие
эпизоды в него поступали воды открытого моря и
проникали нормально морские организмы. Наиболее вероятным источником фосфора были
прилегающие с востока и запада суши. где размы-
вались основные породы.
ДРУГИЕ ВОЗМОЖНЫЕ
МОДЕЛЬНЫЕ ОБЪЕКТЫ
Одним из своеобразных модельных объектов
являются серные и пурпурные бактерии-магнето-
тактики. образующие внутри клеток кристаллы
(домены) магнетита (Kirschvink et al.. 1985: De-
vouard et al., 1998; Postal et al., 1998). Интерес к
ним определяется нахождением похожих струк-
тур в метеорите Оргей (Palik, 1962). Совсем не-
давно были получены аналогичные данные и по
метеориту Мигей.
Вне всякого сомнения перспективны исследо-
вания Fe-бактерий в железистых породах, в том
числе в породах крупных месторождений Ре.
Представляется очень перспективным объек-
том чистые графит и керит. Ранее был исследо-
ван графит из Ботогольского щелочного массива
и было показано вероятное присутствие в графи-
тах биоморфных образований (Жмур и др., 1996).
Очень интригующие результаты были получены
по кернту (см. ниже).
МЕТЕОРИТЫ
На Землю постоянно поступает огромное ко-
личество космического материала. Он поступает
в виде космической пыли и. значительно реже. в
виде достаточно крупных обломков. Оценки ко-
личества внеземного вещества, поступающего на
Землю, весьма противоречивы. Но наиболее при-
нятые - это 100-1000 т в сутки.
По существующим представлениям метеори-
ты имеют астероидальное или кометное проис-
хождение. Все они по составу разделяются на ка-
менные, железокаменные и железные в зависи-
мости от содержания силикатных минералов и
никелистого железа. Подавляющее большинство
обнаруженных метеоритов - каменные, среди ко-
торых также преобладают так называемые хонд-
риты. Каждый из трех упомянутых выше типов
метеоритов подразделяется на классы.
Среди железных метеоритов установлено два
класса: палласиты и мезосидерпты.
Среди каменных метеоритов различают хонд-
риты и ахондриты. Хондриты получили свое на-
звание от хондр - специфических сфероидальных
образований. Среди хондритов выделяется три
класса [энстатитовые (Е). обыкновенные (О) и
углистые (С)] и 9 групп, различающихся по хими-
ческому составу (табл. 1).
Для наших целей наиболее интересными ока-
зываются углистые хондриты. среди которых вы-
делено четыре группы (I. M. О. V). Обозначения
I, M. О. V соответствуют первым буквам типич-
ных метеориов (Ivuna. Mighea. Omans и Vigarano).
Таким образом, например, сочетание СО означа-
ет углистый хондрит типа Omans. Названия мете-
оритов даются по местности.
CI хондриты довольно редки и мелки, и мате-
риалы по ним немногочисленны. В отличие от
всех остальных хондритов. CI хондриты не имеют
настоящих хондр. Основной материал сложен из
мельчайших зерен монтморилонита (глинистый
минерал); септохлорита ∙водный силикат, общая
формула Feft(Si40,n)(OH)└) и магнетита. Иногда
присутствуют в незначительных количествах и
другие минералы.
СМ хондриты распространены значительно
шире. Известные находки достаточно крупные.
Хондриты СМ группы содержат ярко выражен-
ные хондры и непереплавленные агрегаты, по-
груженные в матрицу. Отдельные минералы
представлены оливином, пироксенами. хроми-
том, Са-А1 силикатными стеклами и некоторыми
другими минералами (кальцит, гипс, магнетит). В
матрице наиболее распространены септохлорит
и в меньшей степени монтморилонит. Углерод
СМ хондритов находится в виде сложных углево-
дородов. реже кальцита.
СО хондриты так же. как и СМ хондриты. со-
держат хондры, непереплавленные агрегаты и
одиночные кристаллы и их обломки. Наиболь-
шие различия СМ и СО в содержании хондр (зна-
чительно больше у СО). Хондры могут состоять:
из гранулярного оливина, погруженного в чистое
стекло.низкокальциевых пироксеновых волокон
в стекле, зонарного оливина и железисто-хромис-
той шпннели в стекловатой основной массе. Аг-
регаты неправильной амебовидной формы как
правило сложены оливином с незначительной
примесью пироксена, шпинели и Na-Al-силика-
тов. В матрице были отмечены глинистоподоб-
ные водные силикаты и оливин. Углистая состав-
ляющая в виде сложных углеводородов.
Таблица 1. Химические составы \сндритов (по Р.Т. Додду, 1986. с сокращениями)
|
Класс
|
Эстатптовые (Е)
|
Обыкновенные (О)
|
Углпстые (С)
|
|
Группа %
|
|
|
H
|
L(LL)
|
1
|
M
|
0
|
V
|
|
Si
|
16.47
|
20.48
|
17.08
|
18.67
|
10.40
|
12.96
|
15.75 .
|
15.46
|
|
Ti
|
0.03
|
0.04
|
0.06
|
0.0
|
0,04
|
0.06
|
0.10
|
0.09
|
|
Al
|
0.77
|
1.06
|
1.22
|
1.277
|
0.84
|
1.17
|
1.41
|
1.44
|
|
Cr
|
0.24
|
0.23
|
0.29
|
0.31
|
0.23
|
0.29
|
0.36
|
0.35
|
|
Fe
|
33.15
|
22.17
|
27.81
|
21.64
|
18.67
|
21.56
|
25.82
|
24.28
|
|
Mn
|
0.19
|
0.12
|
0.26
|
0.27
|
0.17 .
|
0.16
|
0.16
|
0.16
|
|
Mg
|
10.40
|
13.84
|
14.10
|
15.01
|
9.6
|
11.72
|
14.52
|
14.13
|
|
Ca
|
1.19
|
0.96
|
1.26
|
1.36
|
1.01
|
1.32
|
1.57
|
1.57
|
|
Na
|
0.75
|
0.67
|
0.64
|
0.70
|
0.55
|
0.42
|
0.46
|
0.38
|
|
К
|
0.09
|
0.05
|
0.08
|
0.09
|
0.05
|
0.06
|
0.10
|
0.03
|
|
P
|
0.30
|
0.15
|
0.15
|
0.15
|
0.14
|
0.13
|
0.11
|
0.13
|
|
Ni
|
1.83
|
1.29
|
1.64
|
1.10
|
1.03
|
1.25
|
1.41
|
1.33
|
|
Co
|
0.08
|
0.09
|
0.09
|
0.06
|
0.05
|
0.06
|
0.08
|
0.08
|
|
S
|
5.78
|
3.19
|
1.91
|
2.19
|
5.92
|
3.38
|
2.01
|
2.14
|
|
H
|
0.12
|
Сл
|
Сл
|
Сл
|
2.08
|
1.42
|
0.09
|
0.38
|
|
С
|
0.43
|
0.84
|
Сл
|
Сл
|
3.61
|
2.30
|
0.31
|
1.08
|

Рис. б. Литифицированные остатки микроорганизмов в метеоритах; а - микроколония коккоидных цианобактерий ти-
па Gloeocapsa l метеорит Мурчнсон : '7 - "ячеистые" остатки по капсулам микроколонцй цианобактерин типа Gloeocap-
sa (метеорит Ефремовка): в - лпп.^о-тетракокки предположительно бактерии Synechococcus (метеорит Ефремовка);
г - остатки гликокаликса. покрывлзшсго коккоидную бактерию, предположительно Aphonoteca (метеорит Ефремов-
ка); д - нитчатая бактерия с клетк:;ми разного диаметра типа Mastigoeladus (метеорит М\рчисон).
CV хондриты - последняя группа, отличается
от СО хондритов более всего по структуре.
Несколько слов о возрасте хондритов. Более
или менее достоверные определения рубидиево-
стронциевым методом были сделаны для групп
обыкновенных и энстатитовых i И, L, LL и Е) хон-
дритов, а также для всех классов свинцово-свин-
цовым методом. Все полученные значения нахо-
дятся в интервале 4.39-4.59 млрд. лет тому назад.
СОВРЕМЕННЫЕ ИССЛЕДОВАНИЯ
МЕТЕОРИТОВ
В последние годы нами в углеродистых метео-
ритах Мурчнсон, Ефремовка. Алленде. Оргей и
др. были выявлены лнтифпцпрованные остатки
коккоидных и нитчатых форм. по морфологии и
размерам аналогичные современным бактери-
альным формам и их древним аналогам (Жмур и
др., 1997; Hoover, 1997: Hoover et al.. 1998). Из вы-
явленных в метеоритах коккоидных форм в пер-
вую очередь укажем на бактериальные формы
из метеорита Мурчисон, близкие по морфоло-
гии к современной колониальной цианобактерий Gloeocapsa. которые, как уже отмечалось
выше, составляли основу цианобактериального
бентосного сообщества, принимавшего участие
в формировании, например, шунгитов Карелии.
Современные макроколонии и микроколонпп
этих организмов часто окружены утолщенной
капсулой, состоящей из структурированных по-
лисахаридов. Эти капсулы, наподобие цист совре-
менных одноклеточных бактерий, несут защит-
ную функцию. Характерной их особенностью яв-
ляется устойчивость при деструкции биомассы и
поэтому они часто сохраняются в ископаемом со-
стоянии. Вероятнее всего, остатки такой капсу-
лы. покрывавшей макроколонню коккоидных
организмов, сохранились в метеорите Мурчисон
(рис. 6. а), сквозь которую просматривается
сложное фрагментарное внутреннее ее строение.
Остатки бактерии типа современной Gloeocap-
sa сохранились и в метеорите Ефремовка. Пред-
ставлены они литифицпрованными капсулами.
окружавшими микроколонии, что определило
"ячеистый" их облик, но в целом сохраняющими
общий первичный облик организма (рис. 6. о\
Напомним, что пример подобной сохранности

Рис. 7. Лнтчфнцированные остатки нитчатых микроорганизмов в метеоритах: а - фоссилизированные остатки нитча-
той циапобактерии типа Micrpcoieus (метеорит Ефремовка); б - нитчатые микроорганизмы с клетчатой фрлгмента-
цией. наподобие современной цнанобактерин Lyngbia (метеорит Алленде I: и - нитчатая бактерия с ветвлением; г -
ветвящиеся нитчатые формы, напоминающие актиномицеты: в, г (метеорит Ефремова).
колоний коккоидных организмов в ископаемом
состоянии был показан на капсулах бывших кле-
ток в современном мате из Себхи Гавиш Синай-
ского полуострова (Gerdes. Krumbem, 1987).
В метеорите Ефремовка были обнаружены
отдельные небольшие по размерам кокковидные
формы, которые имеют тенденцию к группиро-
ванию, причем к такому, которое обычно приоб-
ретают делящиеся клетки, образуя дипло-тетра-
кокки, т.е. состоящие из 2 - ^ клеток (рис. 6, в). Из
современных цианобактерип такое деление наи-
более показательно для представителей Synecho-
coccus. В этом же метеорите были выявлены
ажурные полусферы (рис. 6. г " , вероятнее всего
представляющие литифицированные остатки гли-
кокаликса. покрывавшего коккоидные организ-
мы. Развитие гликокаликса обычно для совре-
менных матов. Он часто сохраняется и после дес-
трукции организмов. Покрытие гликокаликсом в
данном конкретном случае могли иметь различ-
ные коккоидные организмы. Ими могли быть предствители Aphonoteca или пурпурных серобак-
терий Thiocapsa или Thiocystes. Но вполне вероят-
но, что ими могли быть и представители Synecho-
coccus, о чем уже говорилось выше.
В указанных метеоритах встречен достаточно
разнообразный набор литифицированных остат-
ков нитчатых бактериальных форм. К наиболее
представительным из них следует отнести фоссп-
лизированные остатки нитчатых организмов ти-
па современного Microcoleus. объединяющего в
общем чехле несколько трихомов. В достаточно
полной мере сохранившиеся остатки подобным
образом построенного микроорганизма были
встречены в метеорите Ефремовка (рис. 7. а). С
достаточной долей уверенности можно полагать,
что остатки, подобные Microcoleus, были зафик-
сированы и в метеорите Алленде.
Отдельные литифицированные нити могут со-
хранять клеточное строение.
Так в метеорите Алленде была встречена фос-
силизированная нить, диаметр которой преобла-

Рис. 8. Филаме:-:-ы и тороидальные формы гормогонии в метеорите Мурчисон.
дает над длиной клеток, что является диагности-
ческим признаком современных осциллаторие-
вых цианобактерий рода Lyngbya (рис. 7, б).
В метеорите Мурчисон были обнаружены ос-
татки цианобактерни типа Mastigocladus, кото-
рым присущи ветвление и присутствие клеток
различного диаметра (рис. 6. д: ~. в).
Совершенно уникальные образования также в
Мурчисоне были обнаружены Р. Хувером
(Hoover. 1997). Одно из них (рис. S) - это длинная
нить диаметром около 3 мк и протяженностью бо-
лее 100 мк, тесно связанная с породой. Местами на-
блюдаются пережимы и создается впечатление
клеточного строения. Здесь же наблюдаются то-
роидальные формы, которые, возможно, явля-
ются фоссилизированными гормогониями.
Другие формы (рис. 9) представляют собой
пучки тонких трубок, погруженных в единую ма-
трицу (возможно, фоссилизированную слизь -
гликокаликс). Они были первоначально приняты
за остатки низших грибов. Однако, более вероят-
но сравнивать их с современными цианобактери-
ями типа Microcoleus.
В метеорите Ефремовка были выявлены био-
генные структуры, трудно поддающиеся иденти-
фикации, но биогенная природа которых сомнений не вызывает. Имеются в виду тончайшие
ветвящиеся нитчатые формы (рис. 7, г), которые
нами (Жмур и др., 1997) предположительно были
отнесены к актиномицетам, но которые, вполне
вероятно, могут представлять собой и минерали-
зованные следы гликокаликса. так как их строе-
ние мало чем отличается от построения слизис-
той капсулы современного мата.
ОСНОВНЫЕ ТРУДНОСТИ БАКТЕРИАЛЬНО-
ПАЛЕОНТОЛОГИЧЕСКОГО ИЗУЧЕНИЯ
МЕТЕОРИТОВ
Наиболее распространенные возражения про-
тив интерпретации "бноморфных" структур в ме-
теоритах как псевдоморфоз по микроорганизмам
могут быть разбиты на 4 группы.
1) утверждение, что все "биоморфные струк-
туры" являются засорением, которое возникает
после падения метеоритов на Землю.
2) утверждение, что нанометровыи уровень
объектов, продемонстрированный при изучении
марсианского метеорита ALH84001, невозможен
для земных живых объектов.
3) соображение, что мы очень плохо знаем
морфологию абпогенных материалов на наноме-
тровом уровне и поэтому можем столкнуться со
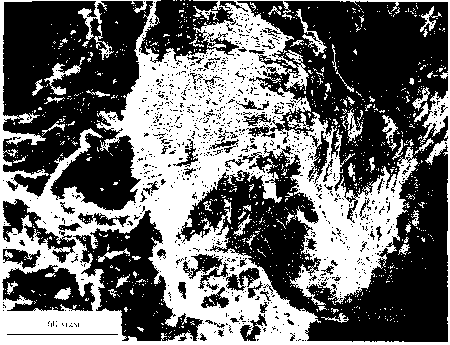
Рис. 9. Фоссилпзировапные остатки, --'ходные с современными цианобактериями типа Microcoleu.s в метеорите М\рчисон
структурами, неотличимыми от тех, которые
принимаются нами за псевдоморфозы по микро-
бам и низшим грибам.
4) утверждение о том, что заведомо абиоген-
ные полимерные кристаллы, например, керита,
не отличимы от фоссилизированных микробных,
а точнее даже цианобактериальных форм.
Эта группа возражений относится прежде все-
го к проблемам морфологического изучения "би-
огенных структур" и является сферой "бактери-
альной палеонтологии".
Остановимся несколько подробнее на этих
возражениях.
1) То. что пребывание метеоритов на Земле
сопряжено с их освоением микроорганизмами -
несомненно. Несомненно и то. что при приготов-
лении препаратов создать исключительно сте-
рильную среду очень трудно. Однако, следует по-
мнить. что мы пытаемся изучать не современные,
не фоссилизированные остатки, а те. которые
уже являются псевдоморфозами, образованными
теми или иными минералами.
Следовательно, если состав исследуемого
объекта не отличается от состава самого метео-
рита. то нет оснований говорить о засорении. Если же отличается, то подозрение о наличии засо-
рения становится очень весомым. Вопрос лишь в
точности химических исследований, например, на
мнкроанализаторах. О многих проблемах выяв-
ления засорений говорилось выше (см. историче-
скую справку).
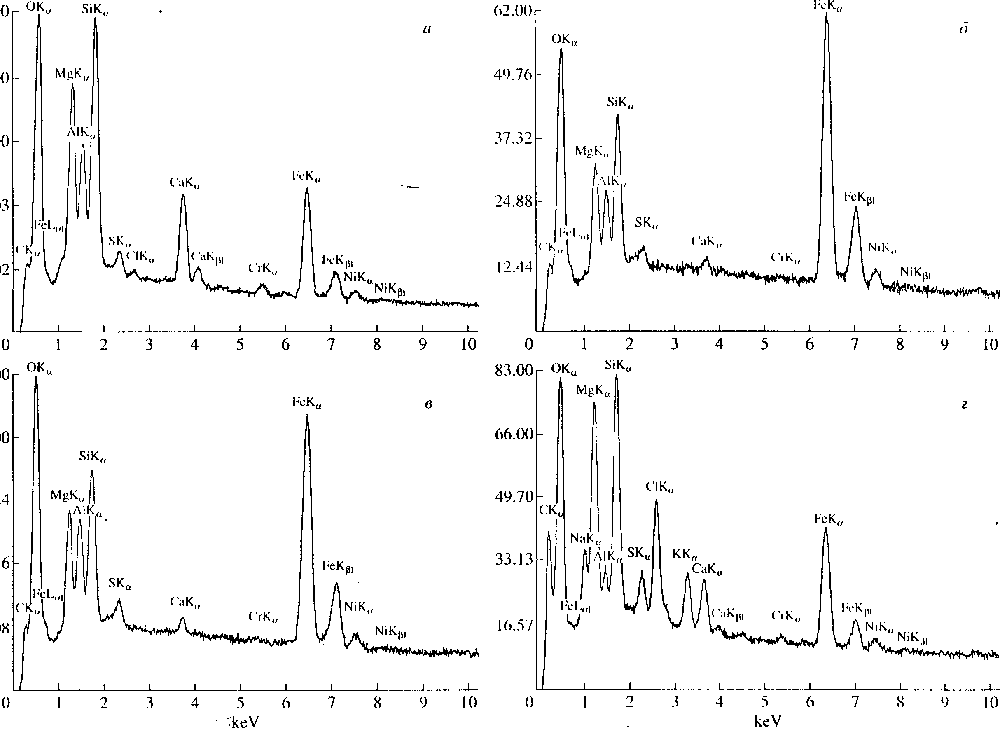
При исследовании метеорита Мурчпсон на
сканирующем микроскопе CamScan с Link были
проанализированы 6 объектов, принятых за псев-
доморфозы по цианобактерням. Сравнение с ре-
зультатами анализа Мурчисона (Додд, 1986), по-
казало, что все проанализированные объекты по
составу не отличаются от матрикса метеорита.
Были также проанализированы на сканирую-
щем микроскопе некоторые объекты из метео-
рита Ефремовка (рис. 10). В тех из них. которые
являются псевдоморфозами, не было обнаруже-
но ничего, выходящего за рамки обычного соста-
ва углистого хондрита. В тех же случаях, когда
было подозрение на засорение, были обнаруже-
ны очень высокие содержания, например. С1 и К.
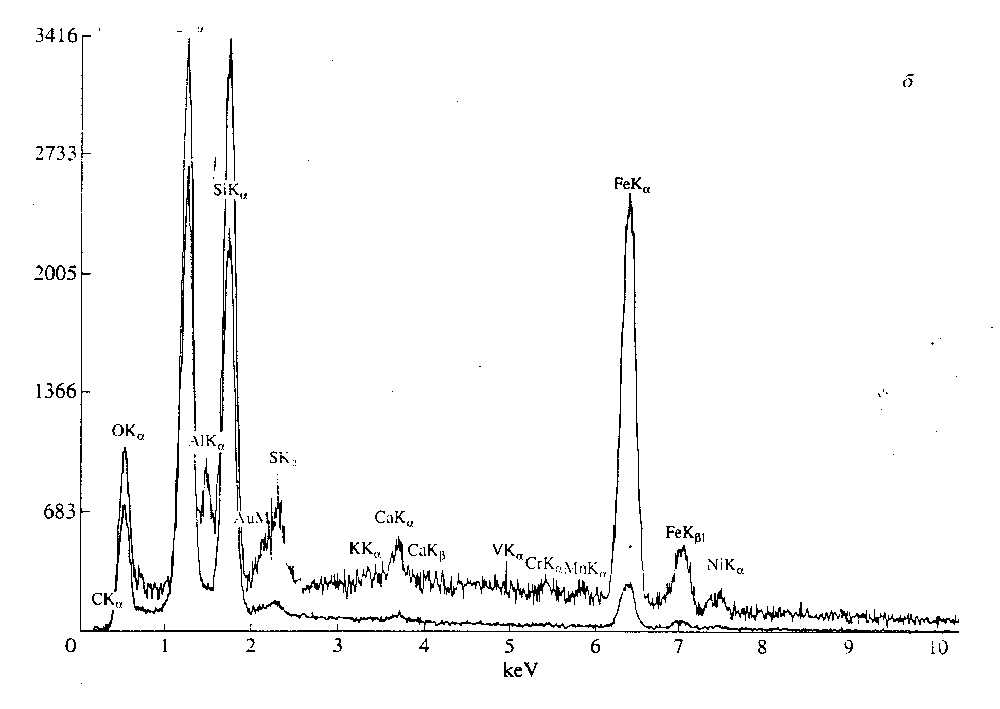
Показательный результат был получен при ис-
следовании метеорита Алленде ( Allende). Один из
объектов при исследовании на ESEM на протяже-
нии короткого времени (несколько минут) менял
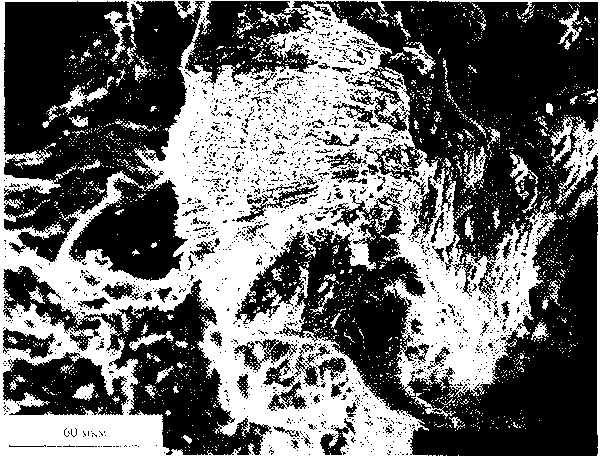
форму (рис. 11. " ). Проделанный анализ выявил
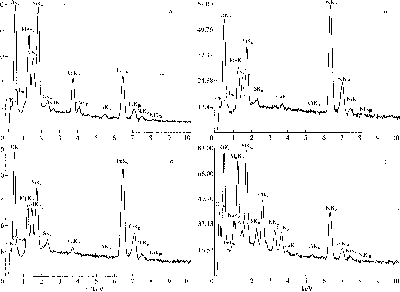
Риг. 10. Графики химических ;|11;|лнчои метеорит;! li(|)peM()iiK;i, полученных ii.i MiiKpoaiiiUiiri.iTope I.ink: ч - м.ггрпкс; i~>, ч - (|)iiJi;iMeirri.i и
iiirrii: / i.icopeime.

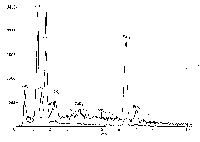
Рис. 11. Метеорит Ллленде: и - фрагмент метеорита, в центральной части расположено горизонтально темно; пятно -
засорение; ii - графики \имичесм'.\ анализов метеорита Ллленде, полученных на микроапализаторе Link. зерхняя
линия - засорение, нижняя - матг.'кс метеорита.
высокие содержания О и Са. но самое главное -
присутствие "значительного количества К и V
(рис. 11,6). Нужно при этом иметь в виду, что ди-
аметр точки, в которой производится определе-
ние. соизмерим с диаметром кокков или нитей.
Поэтому элементный фон породы будет присут-
ствовать почти всегда. Важно для определения
засорений присутствие необычных для метеори-
тов элементов.
Другой аспект проблемы засорений состоит в
том. что при поисках "биоморфных структур" из-
начально мы ищем такие, которые тесно связаны
с породой и не лежат на поверхности исследуемо-
го образца.
Еще раз повторимся: засорения есть и очень
часты. Нередки случаи прорастания метеоритов
актиномицетами или грибами по микротрещи-
нам. Так в образце Мурчисонз из Филд Музея
(Чикаго) было обнаружено, что вся поверхность
образца заражена современными актиномицета-
ми (рис. 12). Но, во-первых, это распознаваемо
приемами, описанными выше. и. во-вторых, это
не может иметь отношения к цианобактериям.
Последние - фотосинтетики, и внедряться внутрь метеорита по трещинам было бы для них "ало-
гичным".
2) Утверждение о невозможности существова-
ния нанометровых микроорганизмов является
очевидным недоразумением. Существует даже
специальное понятие - нанобактерии. Можно
привести множество работ с описанием нанобак-
терии в различных условиях существования и с
описанием их метаболизма (Uvvin.s et al., 1998; Pos-
fai et al., 1998: Kajanderet al.. 1998: Vainshtein et al..
1998: Bjorklund et al└ 1998: Ciftc-ioglu et al.. 1998a. b).
Более того, существуют работы, показывающие
принципиальную неотличимость современных
нанобактерии от объектов из ALH84001 (Folk.
Lynch, 1998).
3) Совершенно иной вопрос о морфологии
абиогенных кристаллов нанометровой размерно-
сти. Думается, что нас могут подстерегать здесь
неожиданности и, несомненно, требуется обоб-
щение уже опубликованного материала, которое.
вероятно, еще не делалось.
Следует сказать, что уже проделаны некото-
рые исследования морфологии нанометровых

Рис. 12. я-г -современные актнномицеты, обрастающие образец метеорита Мурчисон из Филд музея, Чикаго.
кристаллов и полуаморфных выделений фосфатов
из Хубсугульского месторождения (МсКау et al.,
1998). На приведенных фотографиях (рис. 5. в, г)
видно, что обнаружены обычные желваки, веро-
ятно. аморфного фосфата и таблнтчатые крис-
таллы. составляющие трубки цнанобактерий.
Пока нами исследованы апатиты и несомненно,
что дальнейшие исследования должны быть на-
целены в основном на изучение при больших уве-
личениях морфологии тех минералов, которые
встречаются в метеоритах и особенно в углистых
хондритах. Ничего необычного пока не было об-
наружено на нанометровом уровне и при исследо-
ваниях ряда углистых хондритов и марсианских
метеоритов (МсКау et al., 1996".
Вместе с тем иногда среди синтезированных
материалов и. в частности, гетита и гематита на-
блюдаются выделения, весьма напоминающие
бактериальные (Zachara et al.. 1998).
4) Вопрос о морфологии полимерных кристал-
лов действительно очень сложен, и на их природу
существует три разные точки зрения. Уточним,
что речь идет прежде всего о керитах из грани-
тов, точнее камерных пегматитов (Волынь,
Украина), переописанных в последнее время
Н.П. Юшкиным (Yushkin. 1996 . 1998). Состав ке-
рита C.mH^(,0^-S(N). Юшкин приводит следую-
щие параметры формирования кристаллизации
минералов (включая керит): температура - 730-
120╟С. давление - 110-20 МПа. Традиционно при
таких параметрах речь идет исключительно об
абиогенном происхождении таких кристаллов, в
том числе и керита. Однако Юшкин на основании
прежде всего присутствия многих (18) аминокис-
лот и формы кристаллов выдвинул концепцию
"предбиологическнх организмов".
Сходство войлокоподобнои структуры керн-
тов со структурой современных и древних нитча-
тых цианобактериальных матов наводит на мысль,
что исследования керита и вообще графитов
необходимо продолжить, тем более, что нельзя ис-
ключить, что образование керита в упомянутом
случае происходило позднее формирования ми -
нералов на последней стадии гидротермального
процесса.
Кроме возражений "бактериально-палеонто-
логических". существуют возражения по поводу
значения изотопных данных, данных о темпера-
турах формирования ряда минералов, значения
присутствия различных "биомаркеров", в том
числе керогена. пуринов,изопреноидов. циклоал-
ламенов. различных аминокислот и т.п., а также
специфической хиральности аминокислот метео-
ритов. На этих вопросах мы не будем подробно
останавливаться и отошлем читателя к работам,
упомянутым в исторической части (Hodgson, Baker,
1964: Nagy,1975).
Очень часто представления о абиогенном син-
тезе некоторых соединений не доказано, а является результатом явного логического круга. Типи-
чен пример рассуждения очень известного иссле-
дователя геохимии аминокислот Т. В. Дроздовой
(1977, с. 161): "исключением являются лишь угле-
водороды изопренового ряда (фитан и пристан).
происхождение которых связывают в основном с
составной частью хлорофилла - фитолом. Одна-
ко. недавние исследования показали присутствие
нзопреновых углеводородов даже в метеоритах.
что до некоторой степени поколебало наши пред-
ставления о озможности использования этих со-
единений в качестве доказательства связи иссле-
дуемого геологического материала с биогенной
природой этих отложений".
ЗАКЛЮЧЕНИЕ
Значительное морфологическое единство
земных микробных организмов как современ-
ных, так и древних, с псевдоморфозами по микро-
организмам из углеродистых метеоритов, дает
основание полагать принципиальное единство
микробиологического мира Земли и космических
объектов. При этом, конечно, не исключено об-
наружение и таких форм. которые не будут иметь
земных аналогов.
Набор встреченных в метеоритах микрофос-
силий показывает, что это были не отдельные
микроорганизмы, а сообщества, часть из кото-
рых были подобны цнанобактериальному мату.
Эти сообщества, вероятнее всего, принимали уча-
стие в формировании углеродистого вещества ис-
следованных метеоритов.
Вместе с тем, разнообразие организмов, воз-
можно. указывает и на различные обстановки об-
разования микробных сообществ, от термальных
до нормально-бассейновых (типа озерных). Кро-
ме того. следует иметь ввиду, что сравнение с
нанобактериями марсианских метеоритов пред-
полагает весьма аномальные, но допустимые на
Земле условия жизни микроорганизмов (МсКау
et al., 1996).
И. наконец, следует заметить, что исследова-
ния марсианского материала и углистых хондри-
тов шло несколько разными путями. В первом
случае акцент делался на поиск нанобактерий, во
втором пока внимательно исследовался лишь ми-
кронный уровень. Вероятно, использование не-
скольких разных подходов и методик даст значи-
тельный новый материал.
Работа выполнена при поддержке РФФИ.
гранты Х╟ 98-05-64765. .╧ 99-04-48806. .V" 96-05-
97821. ╧97-05-65069.
СПИСОК ЛИТЕРАТУРЫ